Заболевания, связанные с нарушением функции задней доли гипофиза
Вазопрессин и окситоцин - гормоны задней доли гипофиза, правильнее называть нейрогипофизарными гормонами, так как местом образования их является гипоталамус, а именно супраоптическое и паравентрикулярное ядра, где локализуются нейросекреторные нейроны. Аксоны этих магноцеллюлярных нейронов образуют супраоптический гипофизарный тракт, по которому секреторный материал этих нейронов достигает задней доли гипофиза. При микроскопическом исследовании эта часть гипофиза представлена питуицитами, капиллярами, безмиелиновыми волокнами, содержащими нейросекреторные гранулы. Помимо вазопрессина и окситоцина, из задней доли гипофиза выделены соматостатин, тиролиберин и вещество Р.
Иммуноцитохимическими и радиоиммунологическими исследованиями показано, что окситоцин и вазопрессин синтезируются в нейронах супраоптического и паравентрикулярного ядер. Синтезированные в гипоталамических ядрах гормоны транспортируются вместе с соответствующими нейрофизинами по аксонам нейрогипофизарного тракта в заднюю долю гипофиза, где они резервируются "про запас". Транспорт гормонов по аксонам осуществляется в виде гранул, в состав которых входят также специфические белки нейрофизины (белки с мол. м. около 10000). Скорость транспорта гранул гормонов по аксонам составляет 2-3 мм в час. Высвобождение вазопрессина из гранул осуществляется путем экзоцитоза. Вазопрессин может высвобождаться не только в кровь, но и в спинномозговую жидкость, где его концентрация ниже, чем в плазме крови. Не исключается, что секреция гормона в спинномозговую жидкость и заднюю долю гипофиза осуществляется различными нейронами гипоталамуса, т.к. у больных с нейрогенным несахарным диабетом содержание вазопрессина в спинномозговой жидкости в норме.
Однако, помимо основного пути транспорта в заднюю долю гипофиза, у вазопрессина имеется и второй нейросекреторный путь через портальную систему гипофиза к клеткам передней доли гипофиза, где он играет важную роль в регуляции секреции АКТГ. Уровень вазопрессина в портальной системе выше, чем в плазме периферической крови. Это указывает на важность значения вазопрессина в регуляции гипотонии, гиповолемии и других состояний, сопровождающих стрессовую ситуацию.
Ген, ответственный за синтез вазопрессина, локализуется на 20-й хромосоме (20р13). Он содержит три экзона (А, В и С) и два интрона (1 и 2). Ген, кодирующий окситоцин, структурно идентичен описанному, за исключением того, что экзон С короче и кодирует карбоксильный конец нейрофизина и остаток аминокислоты гистидина. Вначале образуется препровазопрессин-нейрофизин II; в процессе транспорта по аксонам происходит отщепление сигнального пептида, образуется прогормон (называемый также пропрессофизином или вазопрессин нейрофизин II), содержащий в своей структуре следующие пептиды: аминотерминал сигнального пептида, вазопрессин, нейрофизин и гликозилированный пептид на карбоксильном конце - копептин. В нейросекреторных гранулах задней доли гипофиза происходит окончательное образование гормона, который по мере надобности высвобождается в центральное кровообращение. Синтез вазопрессина и окситоцина контролируется различными генами, хотя оба локализуются в непосредственной близости друг от друга на 20-й хромосоме (E. Sausville и соавт., 1985).
Нейрофизины (нейрофизин I/НФI/ для окситоцина и нейрофизин II/НФII/ для вазопрессина), которые находятся вместе с гормонами в секреторной грануле, медленно продвигающейся от нейрона до задней доли гипофиза, в течение нескольких лет рассматривались как белки-транспортеры. Однако в дальнейшем было установлено, что они кодируются вместе с вазопрессином (или окситоцином) одним геном и являются составной частью препрогормона, прогормона и высвобождаются в кровообращение одновременно вместе с вазопрессином (или окситоцином), но в свободном состоянии. Нейрофизины биологически неактивны. Время от синтеза гормона и НФ до их высвобождения в системное кровообращение составляет около 1,5 часа. Метаболизм гормонов происходит в основном в печени и почках.
Вазопрессин, или антидиуретический гормон, является основным веществом, регулирующим осмолярность и осмотическое давление жидкостей организма. Выделение и установление химической структуры показало, что у большинства млекопитающих вазопрессин представлен в виде аргинин-вазопрессина, а у свиньи - лизин-вазопрессина, тогда как аминокислотная последовательность окситоцина в молекуле у разных видов одинакова (схема 20).
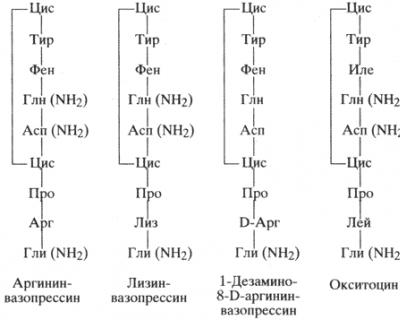
Таким образом, аргинин-вазопрессин, лизин-вазопрессин и окситоцин являются полипептидами, состоящими из 9 аминокислотных остатков, отличающихся друг от друга лишь аминокислотными остатками, расположенными в положениях 3 и 8.
Молекулярная масса вазопрессина составляет 1084, изоэлектрическая точка- рН 10,9; период полураспада - около 10-15 мин. (от 1,1 до 24,1 мин) как у практически здоровых лиц, так и у больных центральным несахарным диабетом и нефрогенным несахарным диабетом. Вазопрессин в циркуляции не связывается белками крови. Однако большая его часть ассоциируется с тромбоцитами, т.к. плазма, богатая тромбоцитами, содержит вазопрессина в 5-6 раз больше, чем плазма без тромбоцитов.
Секреция вазопрессина из нейрогипофизарных нейронов регулируется не только осмо- и барорецепторами, но различными медиаторами: дофамин, серотонин, вещество Р, простагландины, норадреналин, ацетилхолин, ГАМК, глицин, гистамин, ангиотензин II, электролиты плазмы. Синтез и высвобождение вазопрессина стимулируют повышение осмолярности жидкостей организма, гипокалиемия, гипокальциемия, увеличение концентрации натрия в спинномозговой жидкости, снижение артериального давления, уменьшение объема внеклеточной и внутрисосудистой жидкости, стимуляция ренин-ангиотензиновой системы, b-адренергические и холинергические вещества, повышение температуры тела и гипоталамуса, а также ацетилхолин, никотин, апоморфин, морфин (в высоких дозах), адреналин, гистамин, изопротеренол, брадикинин, простагландины, b-эндорфин, винкристин, циклофосфамид, инсулин, 2-деоксиглюкоза, ангиотензин, литий, хлорпропамид и клофибрат. Секреция вазопрессина угнетается при снижении концентрации натрия в спинномозговой жидкости, повышении артериального давления и увеличении объема крови, снижении температуры тела и в области гипоталамуса, приеме антихолинергических веществ, b-адренергических блокаторов, a-адренергических стимуляторов и угнетении ренин-ангиотензиновой системы, а также при приеме алкоголя, глюкокортикоидов, клонидина гидрохлорида, норадреналина, галоперидола, прометазина, оксилорфана, бутофанола, карбамазепина, морфина (низкие дозы), мусцимола и фенитоина.
Основным регулятором секреции вазопрессина является изменение осмотического давления. Осморецепторы, которые преимущественно локализуются в передней части мозга, а именно в переднем гипоталамусе, реагируют на изменение менее чем на 1% осмолярности плазмы. Внегипоталамические печеночные или портальные осморецепторы реагируют, когда осмолярность плазмы отклоняется на 1% и более от нормальных величин.
Исследованиями (C. Thompson и соавт., 1986; J. Davison и соавт.,1987) установлено, что в норме содержание вазопрессина в плазме составляет 2 пг/мл; осмолярность плазмы - 285-287 ммоль/кг, а мочи - 500 ммоль/кг. При увеличении общей жидкости в организме на 1% осмолярность плазмы снижается на 1% (2,8 ммоль/кг), концентрация вазопрессина - до 1 пг/мл и осмолярность мочи - до 250 ммоль/кг. И, наоборот, при уменьшении количества воды в организме на 2% осмолярность плазмы повышается на 2% (5,6 ммоль/кг), уровень вазопрессина также увеличивается с 2 до 4 пг/мл, осмолярность мочи составляет 1000 ммоль/кг. Таким образом, исходя из этих данных, повышение осмолярности плазмы на 1 ммоль/мл должно сопровождаться увеличением концентрации вазопрессина на 0,38 пг/мл и повышением осмолярности плазмы на 100 ммоль/кг.
Помимо осморецепторов, в регуляции осмотического давления принимают участие барорецепторы левого предсердия, каротидного синуса и дуги аорты. Афферентные нервные импульсы из этих зон приводят к ингибированию секреции вазопрессина. И, наоборот, уменьшение силы нервной импульсации от этих рецепторов приводит к повышению высвобождения вазопрессина (P. Norsk, 1989; K. Goetz и соавт., 1991).
Биологическое значение вазопрессина в организме многообразно. Помимо поддержания нормального осмотического давления в организме, он участвует в следующих процессах: а) поддержание артериального давления посредством барорецепторов и прямым влиянием на сосудистую стенку; б) является одним из регуляторов секреции АКТГ; в) увеличивает процесс высвобождения ТТГ из тиротрофов гипофиза; г) увеличивает синтез простагландинов интерстициальными клетками мозгового слоя почек; д) вызывает сокращение мезанглиальных клеток клубочка; е) обладает митогенным эффектом; ж) вызывает агрегацию тромбоцитов и способствует высвобождению факторов коагуляции - фактора Виллебранда, VIIIc фактора и активатора плазминогена тканевого типа и участвует в процессах ЦНС, в частности, процессах памяти.
Это многообразное действие вазопрессина объясняется его взаимодействием с двумя типами плазматических рецепторов. Внутриклеточным медиатором антидиуретического действия вазопрессина в почках является цАМФ. Однако его гликогенолитическое влияние в печени и гладких мышцах сопровождается повышением внутриклеточного кальция. В последующем было показано, что V1 рецепторы опосредуют действие вазопрессина через диацилглицерин и инозитолтрифосфат, а V2 рецепторы-посредством цАМФ. Последний тип рецепторов локализуется на мембранах клеток кортикальной и медуллярной частей собирательных трубок и восходящей части петли Генле. Взаимодействие вазопрессина с V2 рецепторами активирует аденилатциклазу и образование цАМФ, что сопровождается увеличением проницаемости для воды мембраны клеток собирательных трубок и петли Генле. V1 рецепторы выявляются в гепатоцитах, мембранах печени, гладких мышцах сосудов и тромбоцитах. Клетки аденогипофиза содержат рецепторы к окситоцину и вазопрессину, опосредующие их влияние на секрецию АКТГ. Установлено, что в гипофизе также имеются V1 рецепторы, четко отличающиеся от V1 рецепторов печени. V1, V2 и рецепторы к окситоцину характеризуются близкой структурой и относятся к рецепторам, имеющим 7 трансмембранных фрагментов.
Вазопрессин стимулирует синтез PgE, которые ингибируют влияние гормона на активность аденилатциклазы и таким образом снижают антидиуретический эффект вазопрессина. Назначение ингибитора простагландинов (индометацина) потенцирует антидиуретическое действие вазопрессина. Синтез простагландинов стимулируется также ангиотензином, брадикинином, которые несомненно участвуют в модификации действия вазопрессина на почки.
Биологическое действие окситоцина, который, как и вазопрессин, секретируется в гипоталамусе, направлено на стимуляцию сокращения мышц матки и миоэпителиальных клеток, окружающих альвеолы молочной железы, что обеспечавает поступление молока из альвеол в протоки железы. В этих основных органах-мишенях (матка и молочные железы) выявляются рецепторы, связывающие окситоцин. Ионы марганца и магния усиливают процессы взаимодействия окситоцина с соответствующими рецепторами, которые относятся к плазматическим рецепторам, имеющим 7 трансмембранных фрагментов.
Рецепторы у окситоцину, кроме молочных желез, выявляются также в почках, гипоталамусе, гипофизе и некоторых отделах ЦНС, а к вазопрессину - в стенках артерий, дистальных отделах канальцев почек, гипофизе и гипоталамусе.
Механизм действия окситоцина опосредуется аденилатциклазной системой в условиях обязательного присутствия ионов кальция и магния. Определенное место отводится взаимодействию этих гормонов и простагландинов. Так, показано, что физиологический ответ матки на окситоцин зависит от присутствия простагландинов.
Инактивация окситоцина и вазопрессина осуществляется главным образом почками (40%) и печенью (около 50%). Экзогенно введенный вазопрессин частично (10-20%) экскретируется с мочой в биологически активной форме. При почечной форме несахарного диабета вследствие ухудшения связывания вазопрессина с рецепторами количество экскретируемого вазопрессина увеличивается. В процессах инактивации окситоцина определенное место занимает также молочная железа.
Почитайте ещё медицинских статей:
- Плечевая кость Плечевая кость, humerus, длинная трубчатая кость. В ней различают тело и два конца - верхний и нижний. Тело плечевой кости, corpus humeri, в верхнем отделе округло, а в нижнем -
- Несахарный диабет Гипоталамический несахарный диабет - заболевание, характеризующееся жаждой и экскрецией большого количества мочи с низкой относительной плотностью. Развитие заболевания связано с нарушением синтеза, транспортировки и высвобождения вазопрессина. Прием большого количества жидкости
- Мышцы глотки гортань. Иннервация: п... glossopharyngeus. Кровоснабжение: aa.. pharyngeae ascendens palatina ascendens. M.. palalophaiyngeus (см. "Мышцы неба и зева").
Размещена в категории Эндокринная система