Кальцитонин и механизм его действия
Кальцитонин является гипокальциемическим гормоном и секретируется парафолликулярными или С-клетками щитовидной железы, которые отличаются от фолликулярных клеток не только конечным продуктом секреции, но и происхождением.
Парафолликулярные клетки несколько крупнее фолликулярных и содержат большое ядро, митохондрии, хорошо развитый комплекс Гольджи и ряд нежных гранул, содержащих кальцитонин, который высвобождается в ответ на гиперкальциемию.
Парафолликулярные клетки относятся к клеткам APUD-системы, имеющим нервное происхождение (эктодерма нервного гребешка). У человека кальцитонин синтезируется не только в щитовидной железе, но также в вилочковой и околощитовидных железах.
Кальцитонин человека представляет собой полипептид, состоящий из 32 аминокислот с такой последовательностью: Цис-Гли-Асп-Лей-Сер-Тре-Цис-Мет-Лей-Гли-Тре-Тир-Тре-Глн-Асп-Фен-Асн-Лиз-Фен-Гис-Тре-Фен-Про-Глн-Тре-Ала-Илей-Гли-Вал-Гли-Ала-Про-NH2. Мол. м. 3000 дальтон. Период полураспада около 5 мин. Кальцитонин, помимо мономерной формы, может образовывать путем ковалентной связи димерные и, не исключено, полимерные формы гормона, однако биологически активной является только мономерная форма гормона. Было показано, что в процессе трансляции образуется препрокальцитонин и прокальцитонин с мол. м. около 13 кДа. В настоящее время получен кальцитонин человека, крупного рогатого скота, свиньи, овцы и лососевых рыб. Наиболее эффективным (в 10 раз) в биологическом отношении является кальцитонин лососевых рыб по сравнению с кальцитонином человека. Это связано с более длительным периодом полураспада и более длительным существованием гормоно-рецепторного комплекса.
Специфическим стимулятором секреции кальцитонинна является повышение концентрации кальция в крови более 2,25 ммоль/л (9 мг/100 мл). Кроме того, стимуляторами секреции кальцитонина являются катехоламины, осуществляющие свое действие через b-адренергические рецепторы, холецистокинин, глюкагон, гастрин. Глюкагон и катехоламины, взаимодействуя с рецепторами, увеличивают содержание цАМФ, который стимулирует секрецию кальцитонина, так же как и паратгормона, т.е. цАМФ является внутриклеточным медиатором секреции кальцитонина. Кальцитонин метаболизируется в почках, печени и, возможно, в костной ткани.
Биологический эффект кальцитонина проявляется снижением уровня кальция и фосфора в крови, что является следствием влияния кальцитонина на костную ткань и почки. В кости кальцитонин угнетает процессы резорбции как кальция, так и белкового матрикса. Это проявляется снижением экскреции гидроксипролина и содержания кальция в крови. Одновременное уменьшение фосфора в сыворотке крови является результатом снижения мобилизации фосфора из кости и непосредственной стимуляции поглощения фосфора костной тканью. Кальцитонин ингибирует активность и количество остеокластов. Уже через 1 ч после введения кальцитонина уменьшается образование остеокластов из клеток-предшественников. Механизм действия кальцитонина опосредуется цАМФ и активацией протеинкиназ, что сопровождается изменением активности щелочной фосфатазы, пирофосфатазной активности и активности ферментов.
Инфузия кальцитонина человеку или экспериментальным животным приводит к увеличению экскреции кальция, фосфора, натрия, калия и магния. Кальцийурический и фосфоруретический эффект гормона дополняет его действие на костную ткань, приводящее к гипокальциемии и гипофосфатемии. Как в костной ткани, так и в почках взаимодействие кальцитонина с рецепторами приводит к увеличению внутриклеточного медиатора - цАМФ. Если в костной ткани кальцитонин и паратгормон взаимодействуют с рецепторами, локализованными в основном на остеокластах, то в почках эти гормоны комплексируются с рецепторами клеток-мишеней в различных частях нефрона. Рецепторы к кальцитонинну располагаются в восходящей части петли нефрона и дистальных канальцах, тогда как рецепторы к паратгормону находятся в проксимальных канальцах, нисходящей части петли нефрона и дистальных канальцах.
Наряду с паратгормоном и кальцитонином в поддержании фосфорно-кальциевого гомеостаза большое участие принимает витамин D (D-гормон, холекальциферол или витамин D3).
Витамин D3 образуется в коже из 7-дегидрохолестерина под влиянием ультрафиолетового облучения. Однако витамин D3 является неактивным. Некоторые авторы рассматривают его как прогормон, который путем гидроксилирования (25-гидроксилаза) сначала в печени превращается в 25-гидроксихолекальциферол (25-ОНD2), а затем в почках, также путем гидроксилирования (1-гидроксилаза), в 1,25-дигидроксихолекальциферол (1,25-(ОН)2D3. Гидроксилирование в почках может осуществляться и другим путем (24-гидроксилаза) с образованием 24-25-(ОН)2D3. Гидроксилирование витамина D происходит в митохондриях. Необходимо отметить, что витамин D2 (эргокальциферол), содержащийся в растениях, в организме гидроксилируется тем же путем с образованием 1,25-(ОН)2D2, который по биологической активности эквивалентен 1,25-(ОН)2D3. Активирование витамина D в организме представлено на схеме 26.
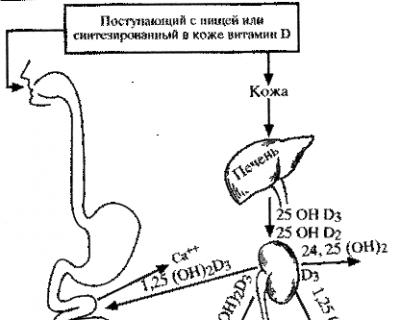
Синтез 1,25-(ОН)2D3 в почках осуществляется при наличии паратгормона и кальцитонина.
Все формы витамина D в организме циркулируют в крови в связанном с белками состоянии. Это a-глобулин с мол.м. около 56000 дальтон. Концентрация витамина D в крови составляет от 1 до 10 нг/мл. Период полураспада около 24 ч. 1,25-(ОН)2D3 действует в кишечнике, увеличивая синтез кальцийсвязывающего белка, ответственного за транспорт кальция через мембрану клеток слизистой оболочки кишечника. В костной ткани 1,25-(ОН)2D3 мобилизует кальций с использованием его вновь образовавшейся костной ткани для процессов минерализации. Это действие витамина не зависит от паратгормона.
Наряду с этим 1,25-(ОН)2D3 влияет на синтез коллагена, участвующего в образовании матрикса костной ткани. Механизм действия витамина в костной ткани блокируется ингибиторами транскрипции и синтеза белка и, вероятно, не отличается от действия 1,25-(ОН)2D3 в кишечнике. 1,25-Дигидроксивитамин D3 оказывает непосредственное влияние на почки, увеличивая реабсорбцию кальция и фосфора в почечных канальцах.
Мышечная слабость, наблюдаемая при недостатке витамине D, объясняется влиянием этого гормона на обмен в мышечной ткани.
Помимо паратгормона, кальцитонина и витамина D, на гомеостаз кальция в организме влияют и другие гормоны.
Почитайте ещё медицинских статей:
- Мышцы спины Мышцы спины, musculi dorsi, делят на поверхностные и глубокие. I. Поверхностные мышцы спины. а) мышцы спины, имеющие отношение к верхней конечности: Трапециевидная мышца, m. trapezius. Широчайшая мышца спины, m. latissimus dorsi. Большая и малая ромбовидные
- Ребра Ребра, costae, числом 12 пар, узкие, различной длины изогнутые костные пластинки, симметрично располагаются по бокам грудного отдела позвоночного столба. В каждом ребре различают более длинную костную часть ребра, os costale,
- Лимфатические сосуды почек и надпочечников Поясничное лимфатическое сплетение, кроме лимфатических сосудов таза и нижней конечности, принимает также лимфатические сосуды почек и надпочечников, поясничной и крестцовой частей позвоночного столба, боковых отделов брюшной стенки и спины. Лимфатические сосуды
Размещена в категории Эндокринная система